Polyphemus (Aa. Th. 1137)
A phylogenetic reconstruction of a prehistoric tale
Julien d’Huy,
Institut Marcel Mauss (UMR 8178) - Équipe LIAS -
École des Hautes Études en Sciences Sociales (France)
http://ehess.academia.edu/JuliendHuy
Abstract: Mythology, like genetics and language, provides essentials elements in the understanding of human history; phylogenetic trees based on mythological versions can allow to reconstruct the history and prehistory of human cultures right back the Palaeolithic period. Indeed, mythological versions are highly conservative. We also can reconstruct the primitive version of a mythological family using phylogenetic reconstructions of ancestral state.
Keywords: mythological structure; mythological evolution; evolution; phylogenetic comparative method; networks; cultural phylogenetics; cultural evolution; ancestral state; vertical transmission; ancestral state; Polyphemus.
Résume: La mythologie, comme la génétique et les langues, fournit des éléments essentiels à la compréhension de l'histoire humaine. Des arbres phylogénétiques basés sur des versions mythologiques peuvent permettre de reconstituer l'histoire et la préhistoire de cultures humaines en remontant jusqu'à la période paléolithique. En effet, les versions mythologiques sont très conservatrices. Nous pouvons également reconstruire la version primitive d'une famille mythologique à l'aide de reconstructions phylogénétiques d'un état ancestral.
Mots clés : structure mythologique; évolution mythologique; évolution; méthode comparative et phylogénétique; réseaux; phylogénétique culturelle; évolution culturelle, état ancestral ; transmission verticale; état ancestral, Polyphème.
Télécharger l'article en pdf / download in pdf: d'Huy - Polyphemus.pdf
Télécharger la base de donnée en doc / download the database in doc: Tableau Polyphemus VERSION DE BASE (1).doc
Cité par / Quoted by:
David Campbell, « Paleolithic Polyphemus : a review », Pleistocene Coalition News,9-1, 2017, p. 6-7
Jérémy Gardent, Mesurer les musiques pour parler du passé. La comparaison des musiques du Gabon comme source d’informations historiques,Thèse de doctorat du Muséum national d’Histoire naturelle, 2017.
Erich Kasten and Michael Dürr, « Jochelson and the Jesup North Pacific Expedition : a new approach in the ethnography of the Russian Far East », inWaldemar Jochelson, The Koryak, 2016, Verlag der Kulturstiftung Sibirien, SEC Publications, p. 9-34.
Patrice Lajoye, « Balor et Yspaddaden Penkawr de par le monde. A propos du motif F571.1 », Nouvelle Mythologie Comparée,2, 2014.
Martine Mussies, Polifemo, MA thesis Musicology, 2013, University of Utrecht.
Jérôme Pace, Mythopoeïa ou l’art de forger les « mythes » dans l’« aire culturelle » syro-mésopotamienne, méditerranéenne et indo-européenne,thèse de doctorat de Sciences religieuses, 2016, Paris, École Pratique des Hautes Études.
Elmas Şahin, « A mythic journey to Polyphemus, Tepegöz and Grendal », International Journal of Literature and Arts,3-2, 2015, p. 12-17.
Adele Tutter, « Embodying disillusionment: Poussin’s blinded giants »,The International Journal of Psychoanalysis,2018, p. 1-27.
Xaverio Ballester, « Mitología galaica primordial », Fol de Veleno, 9, 2020, p. 59-86
Inoslav Bešker, « Brutal Highlanders and Crooked Seasiders », Cross Cultural Studies Review, 1, 1/2, 2019, p. 147-181
Jamshid D. Tehrani, « Descent with Imagination: Oral Traditions as Evolutionary Lineages », in Evolutionary Perspectives on Imaginative Culture, 2020, Springer, p. 273-289.
Benjamin B. Olshin, Lost Knowledge. The Concept of Vanished Technologies and Other Human Histories, 2019, Leiden, Brill
Marc Weiland, Mensch und Erzählung. Helmuth Plessner, Paul Ricœur und die literarische Anthropologie, 2019, J. B. Metzler.
The Finnish school, which has an empirical and positivistic approach, tried to trace, collect and categorize all the variants of a story to reconstruct its history. It tried to establish the pure primeval tale (Urmärchen) from which the versions are originated. Despite the initial enthusiasm, the reconstructive ambitions of the Finnish school have been strongly criticized.
Criticisms of this method can be divided into four main categories: first, it gives a preponderant influence to oral tradition vs. literacy influence; second, the attempt to find the place of origin of tales seems to be doomed to failure, because the evidence has not been evenly collected among countries; third, the tale travelled independently of human migrations; fourth, there is no attempt to effect a reduction that would show how two or more seemingly different themes stand in a transformational relationship to each other.
Recent progresses relating to the development of computational pylogenetic tools, however, may help to resolve the main four problems faced by the Finnish school. First, phylogenetic methods analyze taxa as brothers or cousins, and not in parentage (Levi-Strauss 2002); we also don’t need to presume a gap between the true folktale and literary adaptations. Second, the problem of the origin can be solved by analyzing discrete characters to find the best possible tree. Third, the mythological evolution doesn’t need to follow the same way than the genetic evolution; moreover, computational methods can be used to verify that there is a real tree-like signal in the data. Fourth, there are at least two additional principles (variation and selection) of the folklore transmission being compatible with both evolutionist and structural treatments: the more two myths diverge or transform each other, the more distant is their genetic relationship, geographically and temporally (d'Huy 2012d).
There are many parallels between the process of biological and mythological evolution (d'Huy 2012a, b, d, 2013; see the table) and tools from evolutionary biology are being imported to analyze linguistic and cultural phenomena (Nunn 2011). Additionally, one of the key things about evolutionary science involve reconstructing ancestral states. We also can use phylogenetic comparative methods to make inferences about what thefirst version of a myth was like in the past and how it has changed over time. The main challenge in this research will be the lack of empirical data: prehistoric mythologyleaves almost no traces.
Fig.1.: Myths, like biological species, evolve by a process of descent with modification.
Many parallels can be drawn.
|
GENETIC SYSTEMS |
MYTHS |
|
Discrete heritable units: e.g. the four nucleotides, codons, genes and individual phenotypes. |
Discrete heritable units: e.g. the mythemes, motifs, tale-type. |
|
Mechanisms of replication by transcription and reproduction. |
Imitation, teaching and learning. |
|
Slow rate of evolution. |
Fast or slow rate of evolution. |
|
Parent-offspring, occasionally clonal. |
Parent-offspring, generational transmission, teaching, writing (more recent). |
|
Mutations (e.g. point mutations, insertions, deletions, loss of heterozygosity). |
Innovations (e.g. recastes, variations, mistakes) |
|
Natural selection of traits (individuals who possess certain variants of the trait survive and reproduce more than individuals who possess other variants). |
Social selection of traits (e.g. societal trends, conformity, respect of tradition) |
|
Allopatric or sympatric speciation. |
Geographical or social separation. |
|
Hybridization. |
Mixture between two myths. |
|
Horizontal transmission (e.g. hybridization, transposons and viruses). |
Imposition or extralineal borrowing. |
|
Geographic cline. |
Mythological transformations. |
|
Fossils. |
Ancient myths that have survived in written form. |
|
Extinction. |
Myths death. |
The reconstruction of Polyphemus tale is a textbook case. Many researchers have tried to reconstruct the protomyth, the significance which lies at the root of the story, albeit unsuccessful (for a review, see: Glenn 1978). The most complete attempt for reconstructing the first version of Polyphem was the Hackman's historical and geographical approach (Hackman 1904). Hackman supposed that the original form included i/ the blinding of the giant without his consent, ii/ the flight of the hero under the belly of a ram, iii/ the moment at which the hero gives a false name, iv/ the magic ring episode. According to Hackman, the versions would have travelled to northern Europe via Asia Minor or Greece. Yet Hackman's conclusions were based on statistical criteria that are unclear. Moreover, the author encountered difficulties to arrive at a precise definition of the “original version” of Polyphemus story because of the total lack of explanation for the criteria used to limit the number of versions included into the corpus (Calame 1995: 143). The problem of the physical, geographical origin of the story also seems insoluble (Glenn 1978).
We defined the Polyphemus type as a tale in which a person gets into a homestead – a master of animals' or a monstrous shepherd’s; the host can kill the hero, but he escapes sticking to the hair of an animal who is going out or under an animal’s skin. We tried to solve the specific type of problems we were facing by using powerful statistical tools that are normally used in biology.
We first tested the hypothesis that mythemes, as genes, retained a phylogenetic signature with 79 mythemes in 24 versions of Polyphemus tales belonging to the European and North Amerindian areas (d'Huy 2012c). We used a phylogenetic algorithm, Bio Neigbor Joining (implemented in the program Splitstree 3.2.) to explore global patterns of Polyphemus tales. We obtained a main delta-score of 0.2924. The method scores individual taxa from 0 to 1; a relatively high delta score (close to 1) shows a strong conflicting signal in the data(Holland et al. 2002); it also can be used to estimate the degree of conflict (Gray etal. 2010). The tree obtained agreed with a logical and historical reconstruction of the myth. Moreover, our results showed that the diversification of Polyphem versions was consistent with the distribution of the haplogroup X2. This haplogroup was present in the prehistoric Europe (Lacan et al. 2011, Deguilloux etal. 2011), and has moved from Beringia directly to the North American regions around 18.000 BC, before the Bering land bridge disappeared (Perego et al. 2010).
We decided to go further. We created a new and bigger database including many typological variations of the Polyphemus versions. The data used came from Ojibwa people (Désveaux 1988 : 83), Atsina people (Kroeber 1907: 65-67), Niitsítapi people (Wissler et Duvall 1908: 50-52), Kootenays people (Boas 1918: 213-219, 303-304), Jicarilla Apache people (Goddard 1911 : 21), Kiowa people (Parsons 1929 : 21-26), Lipan Apache (Opler 1940 : 122-125), Homer’s Odyssey (book IX), a medieval French text (Li romans de Dolopathos written by Jean de Haute-Seille), from Oghuz Turks people (Book of Dede Korkut), Berbers (Germain 1935), Palestinian-Israelian people (Patai 1998 : 31-32), Syrian people (Prym et Socin 1881: 115), Serb people (Karadschitsch 1854: 222-225, Krauss 1883: 170-173), modern Greek people (Athens: Drosinis 1884: 170-176; Cappadocia: Dawkins 1916: 551; Chios: Ludwig 1863: 287-289), Albanian people (Comparetti 1875: 308-310), Ossetian people (Dirr 1922 : 262), Abaza people (Colarusso 2002: 200-202; Dumézil 1965: 55-59; ; also named Nart 1 and 2 in the text), Mingrelia people (Frazer 1921: 449-450), Italian people (Abbruzzo : Nino 1883 : 305-307; Sicilian: Crane 1885: 89), Russian people (Ralston 1873: 178-181; Karel 1907: 38-39), Sami people (Poestion 1886: 122-126), Lithuanian people (Richter 1889: 87-89), Hungarian people (Stier 1857: 146-150), Romanian people (Grimm 1857: 15-16), Basque people (Cerquand 1992; Vinson 1883: 42-45; Webster 1879 : 4-6), Gascon people (Bladé 1886 ; Dardy 1884), Swiss people (Abry 2002 : 58), English people (Baring-Gould 1890) and West Highlands people (Campbell 1860 : 105-114).
A structural questionnaire of Polyphemus type was developed on the basis of the known variations in the studied versions. The questions focused on mythemes that are irreducible, unchanging elements in a story. Logically dependent features were eliminated. The mythemes were coded as presence (1) or absence (0) for each version to produce a binary matrix of 98 mythemes in 44 versions. The code incorporated data uncertainty (with the symbol ?).
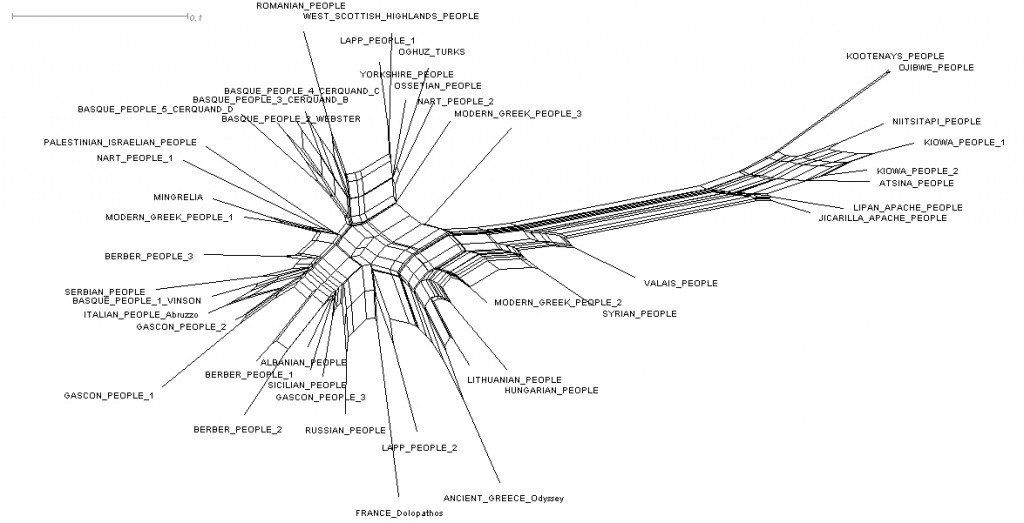
Each mythology borrows mythsfrom neighboring cultures in various contexts.Yet examining network of versions using NeighborNet with Splitstree3.2. reveals a moderate tree-like signal in the tree (fig.2). The resulting network reflects a real (but not dominant) conflicting signal found in structural traits (box-like structures). We must also stress the fact that versions are organized at a very high level with an Eurasian and an Amerindian cluster. Additionally, the graph correctly brings some of the versions together into well-known language or cultural families, with Basque people (Basque people Cerquand B, C, D / Basque people Webster), Gascon people (Gascon 1 and 2), Mediterranean people (first cluster : Greek 1, Berber 3, Serbian ; second cluster with a Berber / Italian group), Black Sea people (first cluster : Mingrelia / Ossetian ; second cluster : Abaza 1 / Oghuz turk), Apache people (Lipan apache and Jicarilla apache) and Algonquian people (Ojibwe / Niitsitapi / Kiowa 1 and 2 / Atsina). Mixed groups could be due to rapid population expansions, conflicting stronger signals (that may be produced by borrowings) and/or non-tree-like descent process (such as hybridization with another myth). Yet these data provide a strong argument in favour of a deep stability of myths.
Fig.2: Splits graphs showing the results of NeighborNet analysis.
The delta score was 0.3639 and the average Q-residual = 0.03148.
The network shows the signal of grouping mythological versions.
The data was then analysed in the phylogenetic package Mesquite 2.75 using a simple model to calculate the parsimony treelength of the tree and matrix (treelength: 334). Then the tree was rearranged by subtree pruning and regrafting. Finally, we rooted the tree between the Ojibwa and the Valais (fig.3).
Fig.3: Parsimony tree rooted with both Ojibwa and Swiss versions.
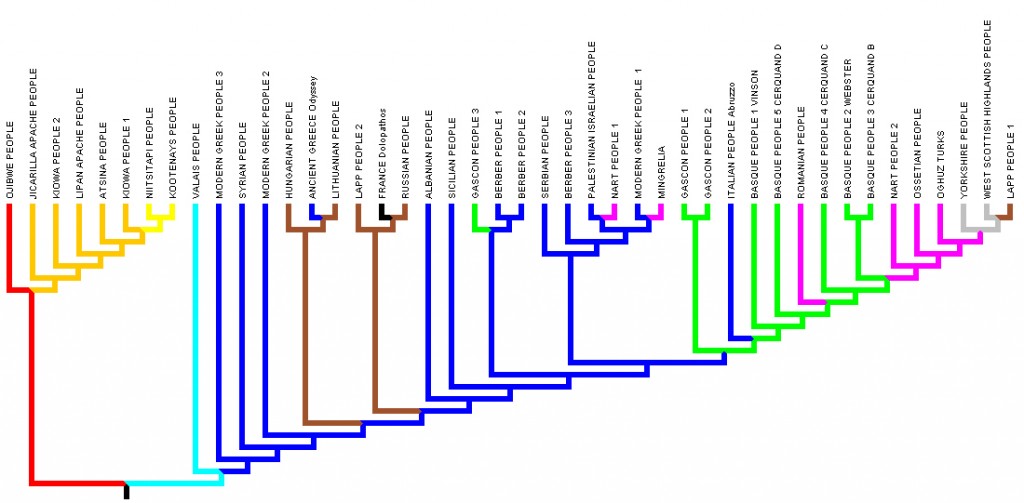
According to the Sapir's age-area hypothesis, the area of greatest divergence in a linguistic family is said to point to the original homeland of the family; when we look at the tree, the greatest divergence is evidently between Ojibwa version and the rest of the family; consequently, the Ojibwa version is the most archaic Amerindian version. The Valais is probably the most archaic European version; it is intermediate between European and Amerindian corpus, with a master of the beasts similar to Amerindian versions. Additionally the closeness in our previous work between Ojibwa and Valais versions corroborates this hypothesis (see d’Huy 2012c).
To check the results, we used MrBayes 3.2.1. to infer the posterior distribution of phylogenetic tree for the 44 versions. We run an ordinary MCMC (Markov Chain Monte Carlo) analysis for 15.000.000 generations. The trees were sample every 1.000 generations. At the end of the run, the average standart deviation of split frequencies was 0,0053. The fact that stationary had been reached was controled with Tracer 1.5.0. Summaries were based on a total of 22.502 samples from 2 runs. Each run produced 15.001 samples of which 11251 samples were included. The tree was rooted with the Valais people version (fig.4).
Fig.4: Consensus tree from the Bayesian analysis of Polyphemus versions.
Node labels show posterior probability (the number is a ratio of one).
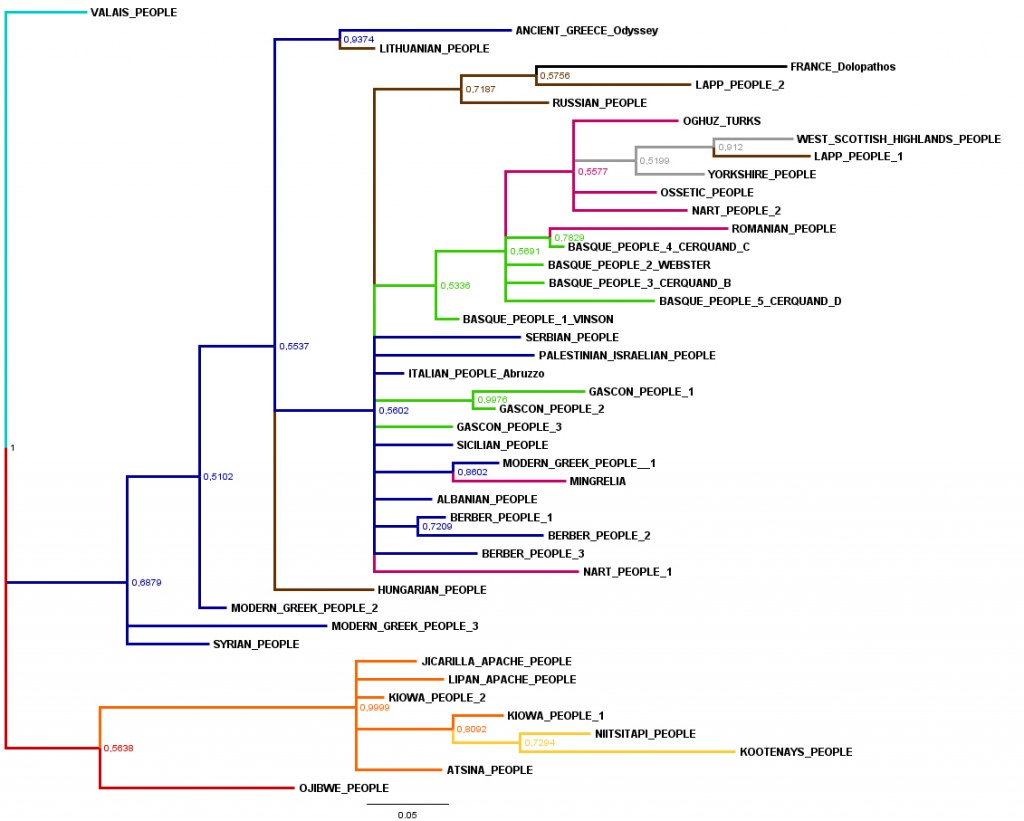
The Retention Index (RI) for the tree calculated with Mesquite 2.75 was 0.612 for the Mesquite tree (fig.3), and 0.792 for the Bayesian tree (fig.4). The RI measures the degree to which each traitis shared by two or more taxa and their most recent common ancestor, whose own ancestor in turn does not possess the trait (synapomorphy). An RI close to 1 is the sign of a tree with a stable and slow evolution; conversaly, an RI close to 0 is the sign of a tree with an unstable or a fast evolution. The both RI we obtain (0.612 / 0.792) are similar or greater than the mean RIs for the biological data sets presented by Lycett and al. (2009), whose main RI is 0.61. The Lycett's biological data sets was probably structured by speciation. So the horizontal transmission (from mother to daughter populations) should be the dominant evolutionary process in both biological and mythological data (see also d’Huy 2012d, e, 2013). Additionally, we obtained a delta-score of 0.3639 for the figure 2. The mean delta-score for European versions was 0.33, and the mean delta-score for Amerindian versions was 0.29. We can conclude that Amerindian mythology is more conservative than the European one.
The trees obtained with Mesquite (fig.3) and MrBayes (fig.4) are not exactly similar. Yet they also are partially consistent with our previous results; the results show a remarkably consistent pattern geographically speaking (North America / Europe; nearest geographical versions tended to form sister clades); the geographical area of the myth is consistent with the haplogroup X2 area in both North America and Europa; Swiss and Ojibwa versions conserve their most archaic place and their proximity can not be explained by a recent areal contact; the closeness between Gascons and Berbers (d'Huy 2012c) is confirmed. On figure 3 and 4, red branches are located in northeastern North America ; orange branches represent the southern region of United States of America ; yellow branches represent the north western regions of United States. Cyan branches represent the Swiss region ; blue branches represent the Mediterranean Sea area ; magenta branches represent the Black Sea area ; Sienna branches are located in northeastern Europe : green branches are located in the Pyrenean area ; grey branches represent the north-western Europe.
According to Malhi and Smith (2002), haplogroup X may have had much more expansive distribution in North America and probably in Europa. The Great Lakes and Near East were probably only a refugium, and not the starting point of initial expansion. So a map that exactly superposed the current haplogroup X diffusion and the Polyphemus versions occurrences don't make sense. Yet haplogroup X is one of the bigger haplogroup among many Amerindian tribes who know the Polyphemus stories, the Algonquian peoples – Ojibwa (25.7%), Niitsítapi (100% but only one sample has been tested), Atsina (11.1%) -, the Kiowa (40%) and the neighboring tribes of Kootenays: the Nuu-Chah-Nulth (11%-13%) and the Yakama (5%) (Smith et al. 1999). Moreover, versions of tribes with haplogroup X are the first branching in the phylogenetic trees, so thse versions probably appear at the beginning of the Amerindian expansion. Strong concentration of haplogroup X2 is also found in the Near East, the Caucasus and Mediterranean Europe; and currently the haplogroup X2 is less strongly present elsewhere in Europe, probably because the last maximum glacial. It is also found among the Berber. Consequently, its expansion agreed with the studied versions area.
Phylogenetic approaches have already provided many insights into the origin and distribution of language and culturally transmitted objects (for a synthesis, see Nunn 2011). Yet, to the best of our knowledge, it is the first time that a geographical distribution pattern allows a phylogenetic reconstitution to go back in time to the Palaeolithic period (to take another mythological instance, see d’Huy 2012d, 2012e).
A historical interpretation of the European branches is problematic. In the first version, conserved in Switzerland, the monster is a master of beasts; this version is largely lost. It could happen during the Late Glacial Maximum when population density in Northern Eurasia decreased. The populations that migrated toward the South (Syria, Greece) probably preserved at least partially the primitive story. Then a new version where the monster was a shelter may be disseminated thanks to successive migrations from the Mediterranean area: with the first migration, the myth was transmitted to northern regions (Lithuanian, Hungarian, then Russian and Lapp); with the second migration, the myth was transmitted to the Gascony ; with the third migration, the myth was transmitted to the Basque area, then in the Black Sea area and in the north-western Europe. Note that links between the Ancient Greek trading cities along the Black Sea cost and the ancestors of the Northwest Caucasian peoples are undoubtedly due to extensive borrowings and influences in the myths of both peoples (Colarusso 2002: 202), e.g. Abaza people 1 and Mingrelia.
The tree with branches proportional to lenghts (fig. 4) shows correlation between total path length from the root of the tree to its tips and number of speciation events (“node”) (fig.5). In other words, where many new mythological versions (“nodes”) appear, there is more total mythological change (longer path lengths). In evolutionary biology, this link indicates a punctuated equilibrium effect (Webster and al. 2003). According to this theory, most species don't change much for the greater part of their geological history; this state is called stasis. When significant evolutionary change occurs, it is generally restricted to rare and very fast events of branching speciation. If correct, newly sister mythological versions tend to diverge rapidly, and are followed by extended periods of stability with little net evolutionary changes. This punctuational effect may thus reflect a human capacity to enhance both the group identity and the identification of individuals with this group (we think that versus they think that)1 or it may be due to a mythological founder effect (d'Huy 2012e). The rest of the time, mythological versions remains inchanged for long period of time, sometimes for thousands of years.
Fig.5: Root-to-tip path length against number of nodes along each path.
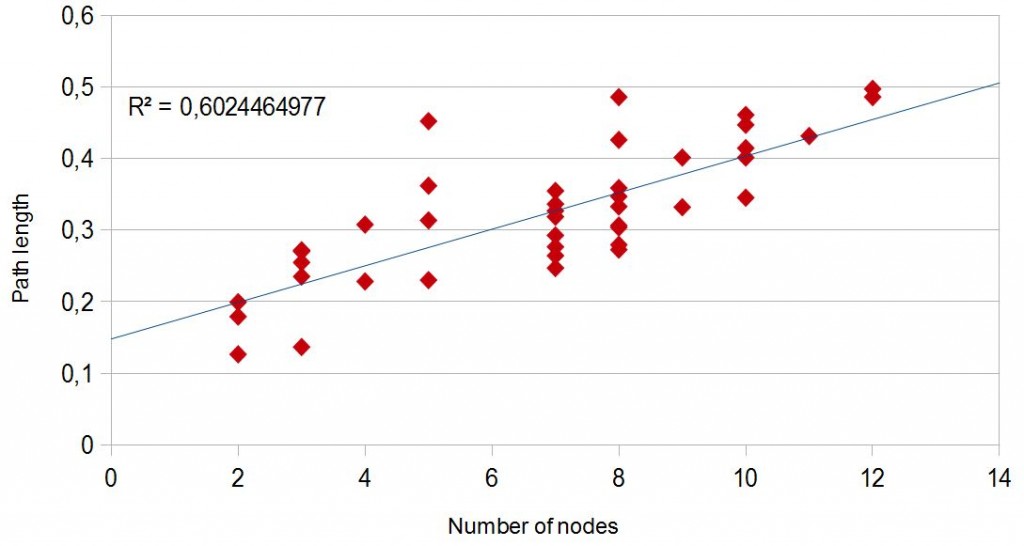
To control the absence of the “node-density artifact” (a well-known artifact of phylogenetic reconstruction that may lead us to believe in a false punctuated equilibrium effect), we calculated the coeficient of determination (R²). An R² near 1.0 indicates that a regression line fits the data well, while an R² closer to 0 indicates a regression line does not fit the data very well. The R² with a linear regression is bigger than the R² with a logarithmic regression. The tree also doesn't seem to show the curvilinear trend that characterizes the node-density artifact (Venditti and al. 2006).
Fig.6: Coeficient of determination of the linear regression (R²)
|
|
linear function |
logarithm |
|
European versions |
0,74 |
0,66 |
|
North Amerindian versions |
0,89 |
0,87 |
|
European and North Amerindian versions |
0,6 |
0,57 |
We used two phylogenetic comparative methods (Maximum Likelihood with model Mk1 and Parcimony reconstructions) implemented in Mesquite 2.75 to examine the probable form of the first state of the Polyphemus family. We applied these phylogenetic reconstruction methods to each mythems of the family. Then we only selected highly confident reconstructed mythems with more than 50% probability with both methods. In the text, we underline mythems with more than 75% probability; the logical connective written in brackets remains just suggestions.
There are at least two monsters; they live in a tent and possess a herd of wild animals. Animals are locked. The hero is a hunter. He enters in the homestead of the monster uninvited, with the express purpose of stealing something, animals or treasure. [Then] the entrance is blocked with a great stone or a locked door. A monster tries to kill the hero and checks the animals that go away. The hero escapes by hiding under the belly of an animal.
This abstract is reconstructed as a simplified version of the Urmärchen. Of course, it is important to remember that the real protomyth probably was as rich in complexity as the versions upon which the reconstruction is based. This protomyth informs us about what Palaeolithic European speakers saying. For instance, it documents evidence for a belief in a master of beasts.
The phylogenetic models used in this paper allowed us to: make accurate inferences about human migration (our data independently corroborate a common and prehistoric origin of Amerindian and European haplogroup X2) taking into account the probability; test the impact of mythological borrowings between neighbouring tribes and the part of similarities appearing by chance; show that mythology evolves in punctuational burst; reconstruct palaeolithic states of a mythological family with quantification of confidence .
The author thanks Jean Peyresblanques and Patrice Lajoye for their help.
Abry C., « Omer m’a tuer… ou moi-même en personne », in : A. Hurst et F. Létoublon [éd.], La Mythologie et l’Odyssée : hommage à Gabriel Germain. Actes du colloque international de Grenoble 20-22 mai 1999, Genève, Droz, 2002, pp.57-65.
Baring-Gould S. (1890), "The Giant of New Mills, Sessay," Folk-lore, I. : 130-131.
Bladé J.-F., Contes de Gascogne, 1 : Contes épiques, Paris, Maisonneuve et Ch. Leclerc, 1886.
Boas F., Kutenai Tales, Washington, Smithsonian Institution, Bureau of American Ethnology, Bull. 59, 1918.
Calame C. (1995). The Craft of Poetic Speech in Ancient Greece. Trad. J. Orion, Cornell University Press, 220 p.
Campbell John-Francis (1860). Popular Tales of the West Highlands orally collected, I. Edinburgh : Edmonston and Douglas,
Cerquand Jean-François (1992). Légendes et récits populaires du Pays Basque. Bordeaux : Aubéron, 347 p.
Colarusso John (2002), Nart Sagas from the Caucasus; from the Circassians, Abazas, Abkhaz and Ubykhs. Princeton and Oxford, Princeton University Press, 552 p.
Comparetti Domenico (1875). Novelline popolari Italiam. Rome, Turin et Florence : Ermanno Loescher, 310 p.
Crane Thomas Frederick (1885). Italian Popular Tales. London: Macmillan, 389 p.
Dardy L., Anthologie populaire de l'Albret, sud-ouest de l'Agenais ou Gascogne landaise, II, Contes populaires, Agen, Michel et Médan, 1891, 2 vols.
Dawkins Richard McGillivray (1916). Modern Greek in Asia Minor: a study of the dialects of Siĺli, Cappadocia and Phárasa, with grammar, texts, translations and glossary. Cambridge: University Press, 695p.
Deguilloux, M.-F., Soler, L., Pemonge, M.-H., Scarre, C., Joussaume, R., & Laporte, L. (2011). « News from the west: Ancient DNA from a French megalithic burial chamber. »American Journal of Physical Anthropology, 144(1), 108-118.
Désveaux E., Sous le Signe de l’Ours. Mythes et Temporalité chez les Ojibwa septentrionaux, Paris, La Maison des Sciences de l’Homme, 1988.
Dirr Adolphe (1922). Kaukasische Märchen. Iena : verlegt bei E. Diederichs, 294p.
Drosinis Georgios (1884). Ländlische Briefe von Georgios Drosinis : Land und Leute in Nord-Euböa, Deutsche Uebersetzung von Aug. Boltz, transl. A. Boltz, Leipzig : Friedrich, 180p.
Dumézil Georges (1065). Le Livre des Héros. Légendes sur les Nartes. Paris : Gallimard, 264p.
Frazer James George (1921). Apollodorus : The Library II. Londres : William Heinemann / New York : G.P. Putnam's sons, 546p.
Germain G., « Ulysse, lecyclope et les Berbères », Revue de Littérature Comparée, n°15, 1935, pp.573-623.
Glenn Justin (1978). « The Polyphemus Myth : Its origin and interpretation. » Greece 1 Rome, vol. 25, no 2 : 141-155.
Goddard P.E., Jicarilla Apache Texts. Anthropological Papers of the American Museum of Natural History, vol. 8. 1911.
Goddard Pliny Earle (1904). Hupa Texts. Berkeley : The University Press, 278p.
Gray R.D., Bryant D. et Greenhill S.J., « On the shape and fabric of human history »,Philosophical Transactions of the Royal Society,365, 2010, pp. 3923-3933.
Grimm W., « Le Mythe de Polyphème », Revue Germanique, vol.IX, 1860, pp.589-648.
Grimm Wilhelm (1857). Die Sage von Polyphem. Berlin, Gedruckt in der Druckerei der Königl . Akademie der Wissenschaften, 39p.
Hackman O. (1904). Die Polyphemsage in der Volksüberlieferung. Helsinki
Holland B. R., Huber K. T., Dress A. and Moulton V. (2002). δ plots : a tool for analyzing phylogenetic distance data. Molecular Biology and Evolution 19 : 2051-2059.
d’Huy J. (2012a). « Le motif de Pygmalion : origine afrasienne et diffusion en Afrique. »Sahara 23, 49-58.
d’Huy J. (2012b). « Mythes, langues et génétique. » Mythologie française, 247, 25-26.
d’Huy J. (2012c). « Le conte-type de Polyphème : essai de reconstitution phylogénétique. » Mythologie française, 248 : 47-59.
d’Huy J. (2012d). « Un ours dans les étoiles: recherche phylogénétique sur un mythe préhistorique. » Préhistoire du Sud-Ouest, 20, to be published.
d'Huy J. (2012e). “A big camel in the Berber sky.” Les Cahiers de l'AARS, 16, to be published.
d'Huy J. (2013). “A phylogenetic approach of mythology and its archaeological consequences.” Rock Art Research 30(1), to be published.
Karadschitsch Won Stephanowissel, with the collaboration of W. Tochter (1854). Volksmärchen der Serben, Berlin,
Karel Jaronomir Erben (1907). Russian and Bulgarian Folk-lore Stories. transl. W.W. Strickland, London : G. Standring, 132p.
Krauss Friedrich S. (1883). Sagen und Märchen der Südslaven in ihrem Verhältnis zu den Sagen und Märchen der übrigen indogermanischen Völkergruppen. Leipsic, W. Friedrich, Vol. I.
Kroeber A.L., « Gros Ventre myths and tales », Anthropological Papers of the American Museum of Natural History 1 (part 3), 1907, pp.55-139.
Lacan, M., Keyser, C., Ricaut, F.-X., Brucato, N., Duranthon, F., Guilaine, J., Crubézy, E., & Ludes, B. (2011). Ancient DNA reveals male diffusion through the Neolithic Mediterranean route. Proceedings of the National Academy of Sciences of the United States of America, 108(24), 9788-9879.
Lévi-Strauss C.- « CR : Guillaume Lecointe & Hervé Le Guyader, Classification phylogénétique du vivant », L’Homme,162, 2002, pp. 309-312.
Ludwig Ross (1863), Erinnerungen und Mittheilungen aus Griechenland. Berlin: R. Gaertner, 313p.
Lycett S.J., Collard M. et McGrew W.C. (2009), « Cladistic analyses of behavioural variation in wild Pan troglodytes : exploring the chimpanzee culture hypothesis », Journal of Human Evolution, 57, pp.337-349.
Malhi Ripan S. and Smith David Glenn (2002). “Brief Communication: Haplogroup X confirmed in Prehistoric North America.” American Journal of Physical Anthropology 119: 84-86.
Nino, Antonio de (1883). Usi e Costumi Abruzzesi III. Florence : Tipographia di G. Barbera, 278p.
Nunn Charles L. (2011). The Comparative Approach in Evolutionary Anthropology and Biology. USA: The University of Chicago Press, 379p.
Opler M.E. Myths and Legends of the Lipan Apache Indians, New York, Memoires of the American Folklore Society, vol.36., 1940 (New York, Kraus reprinting Co., 1969).
Parsons E.C.. Kiowa Tales, New York, Memoires of the American Folklore Society, vol.22., 1929 (New York, 1969: Kraus reprinting Co.).
Patai Raphael (1998). Arab Folktales from Palestine and Israel. Wayne State University Press, 270p.
Perego U.A., Achilli A., Angerhofer N., Accetturo M., Pala M., Olivieri A., Kashani B.H., Ritchie K.H., Scozzari R., Kong Q.P., Myres N.M., Salas A., Semino O., Bandelt H.J., Woodward S.R., Torroni A. (2010) « Distinctive Paleo-Indian Migration Routes from Beringia Marked by Two Rare mtDNA Haplogroups », Current Biology, vol.19, pp.1-8.
Poestion J.C. (1886). Lappländische Märchen, Volkssagen, Räthsel und Sprichwörter. Vienna: C. Gerold, 274p.
Prym Eugen and Socin Albert (1881). Syrische Sagen and Maerchen aus dem Volksmunde . Göttingen : Vandenhoeck und Ruprecht.
Ralston, William Ralston Shedden (1873). Russian Folk-tales. London : Smith, Elder and C°, 382p.
Richter Fr von (1889) "Litauische Märchen. Der einäugige Riese." Zeitschrift für Volkskunde, I.: 87-93.
Smith David Glenn, Malhi Ripan S., Eshleman Jason J., Lorenz Joseph G. and Kaestle Frederika A. (1999). “Distribution of mtDNA Haplogroup X Among Native North Americans.” American Journal of Physical Anthropology 110:271–284.
Stier Gaals Georg Von (1857). Ungarische Volksmärchen. Pesth: Heckenast.
Venditti Chris, Andrew Meade and Mark Pagel (2006). "Detecting the node-density artifact in phylogeny reconstruction." Systematic biology 55 (4) : 637-643.
Vinson Julien (1967). Le Folk-lore du pays Basque. Paris: Maisonneuve, 396p.
Webster W. (1879). Basque Legends, with an essay on the Basque language by M. Julien Vinson, Londres, Griffith and Farran, 1879.
Wissler C. et Duvall D.C., Mythology of the Blackfoot Indians, New York, American Museum of Natural History (Anthropological Papers of the American Museum of Natural History. Vol. 2, Part 1), 1908.
Webster Andrea J., Payne Robert J. H. and Pagel Mark (2003) Molecular Phylogenies Link Rates of Evolution and Speciation. Science 301: 478.
1. For instance, a story of the origin of fire can be told by an amerindian to offset another story by an indian of another tribe (Goddard 1904: 197).
Commentaires
Cela semble très intéressant mais c'est en langue étrangère. Une version dans notre langue est-elle prévue ?
Ma foi, s'il se trouve un volontaire pour traduire l'article, pourquoi pas. Mais pour ce qui concerne les publications scientifiques, qui plus est en science de l'homme, les articles sont pris dans la langue dans laquelle ils sont reçus et dans laquelle ils ont été évalués par le comité de rédaction. Surtout en mythologie, domaine dans lequel tout chercheur se doit d'avoir dans son arsenal une bonne palette linguistique, autrement deux ou trois langues en plus de sa langue maternelle.